


















Allevare api senza dover trattare contro la Varroa Destructor: utopia o realtà?
di Reto Mordasini
Quanto segue è il lavoro finale dell'autore per il conseguimento del diploma svizzero di apicoltura. È stato scritto nel 2019, e da allora molto è stato scritto e molto è stato fatto su questo tema. Il testo è presentato così come scritto allora, salvo qualche aggiustamento stilistico ma senza aggiornamenti, sui quali magari si potrà tornare in futuro
Qui sono inclusi solo l'Introduzione e il primo capitolo, che colloca l'approccio più 'naturale' dell'autore nel contesto delle receni discussîoni teoriche sulla relazione tra gestione zootecnica delle api e la pressione selettiva esercitata dall'ambiente. La seconda parte presenta l'approccio concreto dell'autore nel suo percorso di valorizzazione della selezione naturale; decideremo in seguito in quale forma rendere noti questa procedura e i suoi risultati.
Indice
1.1 L’ectoparassita Varroa e i virus associati
1.1.2 La trasmissione della varroa
1.1.3 I virus associati alla varroa
1.2 Difesa immunitaria dell’ape.
1.2.3 Immunità sociale delle api e varroa (VSH)
1.3.1 L’apicoltura e la selezione naturale.
1.3.2 Selezione naturale e l’apiario.
1.3.3 Selezione naturale e l’alimentazione artificiale.
Introduzione
L'acaro ectoparassita varroa Destructor e i virus patogeni ad esso associati sono una minaccia per le colonie di api, e gli apicoltori sono costretti a controllare il parassita (trattamenti) per mantenere in vita le proprie colonie. Ciò nonostante, ogni anno vi sono delle perdite anche numerose di famiglie di api da miele.
Tuttavia il problema varroa non è solo un rapporto tra api e acari, ma è un evento evolutivo in cui anche i virus, l’habitat e gli apicoltori sono direttamente coinvolti.
Conoscere il vettore varroa è stato di primaria importanza: mi ha permesso di capire come le pratiche nella gestione dell’apiario possano influenzare una pressione evolutiva su ape, acaro e virus, così da comprendere al meglio le dinamiche che li legano. Il problema che mi sono posto è stato dunque quello di capire cosa ricercare nei comportamenti delle api in relazione all’igiene immunitaria nei confronti della varroa, e come valutare il grado di infestazione del parassita nelle famiglie di api, in modo di riconoscere e non diffondere genotipi di api “non resistenti” che dipendono da trattamenti ripetuti.
Nel corso degli anni sono riuscito a diminuire sensibilmente il ricorso agli acaricidi e agli interventi biomeccanici contro la varroa, non solo in quanto tossici anche per le api o per il fatto che rompano un equilibrio naturale, ma anche in quanto vanno ad influenzare il sistema immunitario individuale e sociale delle api.
In natura, questo ha dato alle api la possibilità di sviluppare naturalmente meccanismi di resistenza in risposta alla pressione selettiva degli acari varroa. Con il tempo sono riuscito a identificare e selezionare api che presentavano caratteri “speciali” riconducibili ai tratti Varroa Sensitive Hygiene (VSH). Queste famiglie di api selezionate, riescono a mantenere il grado d’infestazione di varroa basso nell’arco della stagione attiva, così dover ricorrere ad un unico trattamento annuo. Dopo l’unico il trattamento in tardo autunno, le famiglie, hanno una caduta tra 96 e i 650 acari per arnia e poche api in poche colonie presentano segni particolari di virus (DWV). Le mie perdite invernali a causa di varroasi o virus associati sono molto contenute, a dipendenza dell’annata e dalla posizione geografica. Ho invece delle perdite legate alla carenza di cibo, in quanto la mia apicoltura è fortemente improntata ad una selezione naturale ed evita nel limite del possibile l’introduzione nell’arnia di qualsiasi cosa non prodotta dalle api stesse.
La mia apicoltura è in continua metamorfosi e negli ultimi 10 anni è passata da “attiva” ad essere sempre più “passiva”; alla sua base vi sono l’osservazione dell’alveare e il benessere psicofisico del superorganismo ape.
Non sono ancora riuscito a gestire api senza alcun trattamento, ma da un paio di anni la mia pratica apistica si è notevolmente alleggerita, per me come apicoltore (minor manodopera e minor investimento finanziario), ma soprattutto minor stress per le api.
La prima parte di questo lavoro è basata sui lavori scientifici degli autori che hanno studiato questi argomenti, e costituisce le fondamenta delle procedure che ho adottato e che descrivo nella seconda parte. Quest’ultima è il risultato di esperienze personali, ma ritengo comunque che i risultati siano incoraggianti.
Capitolo 1: l'evoluzione del sistema ape-varroa
1.1 L’ectoparassita Varroa e i virus associati
1.1.1 La Varroa Destructor (Anderson & Trueman, 2000)
L'obiettivo di questa parte è conoscere meglio la varroa per meglio comprendere come le pratiche di gestione dell’apiario possano influenzare una pressione evolutiva sull’ape, sull’acaro e sui virus.
L’ acaro Varroa destructor è originario dell’Asia, dove convive con il suo ospitante nativo (Apis cerana) che, grazie alla coevoluzione, è riuscito a tenere sotto controllo il parassita, sviluppando delle tecniche, come l’autosacrificio delle larve femminili, oppure con la “sepoltura” della cella opercolata maschili con uno strato di cera particolarmente tenace. In questo modo blocca lo sfarfallamento del fuco causandone la morte e di conseguenza la varroa e la sua prole viene imprigionata in una cella fatale senza via d’uscita (Peng)1. In tutte e due i casi l’Apis cerana “disturba” il suo parassita durante il periodo più vulnerabile, la riproduzione, così da non permettere alle nuove generazioni di acari di continuare il proprio ciclo vitale.
La varroa é un parassita cieco, indifeso, che si affida esclusivamente al tatto, forse al gusto ma soprattutto all’olfatto, poiché gli acari sono in grado di riconoscere diverse miscele di idrocarburi sulla cuticola delle api, in modo di riconoscere e selezionare le api nelle loro varie fasi di vita (nutrici, bottinatrici, vari stadi larvali, ecc.) o scovare api appartenenti a colonie diverse (Del Piccolo)2.
La varroa dipende in tutto e per tutto dalle api, per gli spostamenti, per la termoregolazione, per la riproduzione e per la nutrizione, che ottiene utilizzando parti della bocca specializzate per perforare la cuticola dell'ape adulta o la larva in via di sviluppo (Sumpter, Martin)3. L’acaro non si nutre di emolinfa, ma danneggia le api e le pupe ospitanti, consumando il loro corpo grasso (vitellogenina), un tessuto che dovrebbe restare integro, per una corretta funzione immunitaria, come riserva alimentare, ha funzioni disintossicanti e anche altri processi essenziali per la salute delle api (Ramsey, Bauchan, Gulbronson)4.
Ogni fase del ciclo vitale della varroa dipende dalla sua capacità di posizionarsi in modo sicuro e di rilevare piccoli dettagli e differenze chimiche che le consentono di sincronizzare i tempi riproduttivi con lo sviluppo della larva ospite. Per sopravvivere, l’acaro ha sviluppato strategie sofisticate per adattarsi al parassitato, come l’addome concavo, così da potersi aggrappare saldamente al corpo dell’ape.
La varroa é in grado di assimilare i ferormoni della famiglia di api ospitante, grazie allo strato lipidi di cui è ricoperto il suo esoscheletro ed in sole tre ore farsi invisibile, diventando un fantasma nel buio dell’alveare (Kather)5.
I delicati acari immaturi muoiono rapidamente al fuori di una cella opercolata, ma anche una femmina matura può morire al morso di un'ape.
La varroa, è arrivata in Europa negli anni ottanta, ma l’ape Mellifera e gli apicoltori si trovarono impreparati a questo neobiota parassita invasivo che causava la morte degli alveari. Così, come di prassi, l’industria agricola ha impiegato i pesticidi per combattere l’infestane acaro, il quale è stato in grado di evolvere rapidamente grazie al suo ciclo generazionale di soli 17 giorni e adattarsi alle nuove situazioni, così da diventare immune a vari acaricidi impiegati. Invece, l’ape europea, che ha un ciclo generazionale di circa un anno, non ha avuto il tempo di adeguarsi ed evolversi, anche a causa delle pratiche apistiche che rimuovono l'acaro, e di conseguenza non è stata soggetta alle pressioni selettive richieste per tale processo.
1.1.2 La trasmissione della varroa
La “trasmissione” della varroa avviene per lo più per via orizzontale, cioè da ape in ape e da arnia a arnia, più delle volte dovuto al saccheggio di famiglie deboli o morte, oppure dalla deriva di api parassitate indotta dalla concentrazione di colonie di api che vivono ammassate in un apiario in maniera del tutto innaturale. In piccola parte la trasmissione dell’acaro avviene per via verticale con la sciamatura.
In natura, le colonie sono spesso ampiamente distanziate, in base alla disponibilità di cavità per nidificare e alla disponibilità di foraggio del territorio. Seeley T. contò una densità di circa 2,5 colonie di Apis Mellifera per miglio quadrato nella foresta selvaggia di Arnot (Seeley)6. In queste condizioni, è difficile che una bottinatrice non faccia ritorno al proprio alveare andando alla deriva.
Mentre in alveari disposti in file, a distanza di un metro e dipinti dello stesso colore e rivolti nella stessa direzione, è comune che il 40% o più di tutte le api operaie si spostino dalla loro colonia natale a una colonia vicina e ciò fa aumentare il rischio di acquisire patogeni e parassiti dalle colonie vicine (Seeley)7.
La questione della deriva presenta due spetti. Da una parte, mentre il 70% delle api infestate da varroa inizialmente si orientate verso l'ingresso sbagliato, le api contagiate da acari avevano il doppio delle probabilità di ritornare ad un'entrata adiacente "fittizia" piuttosto che all'ingresso corretto del loro alveare8. Dall’altra, si è dimostrato che l’abbondanza di varroa, influisce sui profili chimici delle api operaie, e che colonie infestate da varroa non sono più inclini alla deriva, ma piuttosto hanno permesso l’ingresso di api di un’altra famiglia (Cervo)9.
1.1.3 I virus associati alla varroa
La colonia di Apis mellifera, l'acaro ectoparassita Varroa e i virus associati, formano congiuntamente un complesso sistema di interazioni ospite-parassita.
In un primo momento, sembrava che la causa maggiore di perdite delle colonie fosse il parassita stesso, ma in realtà la varroa si è rilevata solo il vettore di virus, dal momento che una colonia muore tipicamente per un'epidemia di uno o più virus trasmessi dal parassita, molto prima che soccomba per l'impatto dell'acaro stesso (Genersch, von der Ohe)10.
È stato dimostrato che i virus della paralisi acuta (APV), i virus delle ali deformi (DWV), i virus della paralisi lenta (SBPV) e il virus del Kashmir (KBV) possono essere trasmessi con successo alle api durante le attività di alimentazione degli acari.
Le attività di alimentazione dell'acaro varroa ha favorito una nuova via di trasmissione per alcuni virus delle api, precedentemente trovati solo in forma inapparente e la trasmissione era esclusivamente attraverso le vie oro-fecali e che le api potevano generalmente tenere sotto controllo grazie alla forte immunità alle infezioni, date dal loro intestino.
Almeno 14 piccoli (17-60 nm) virus RNA sono noti sull'ape mellifera e sono normalmente presenti nella maggior parte delle colonie apparentemente sane, sotto forma di una comune infezione inapparente con basso livello di particelle virali. (Sumpter, Martin)11.
Queste infezioni asintomatiche rendono la definizione di salute più complessa, in particolare nelle api sociali, in cui individui di diverse caste, età e background genetici (cioè, che variano in relazione con la poliandria) formano un ‘superorganismo’ (Engel)12.
L'insorgenza di deformità fisiche nell’ape è associata alla trasmissione del virus DWV per mezzo della varroa durante gli stadi pupali dell’ape. Il virus DWV è un patogeno virale dell'ape che causa deformità alare ed è una delle cause maggiori di morte nei nostri alveari.
Le analisi effettuate sulle api che palesano una conclamata infezione di DWV, rivelano che quasi tutte le parti del corpo erano positive al virus DWV (testa, il torace e l'addome). Al contrario, per le api asintomatiche, le sequenze virali RNA sono rilevate unicamente nel torace e/o nell’addome, ma mai nel RNA estratto dalla testa. Il virus DWV è anche stato rilevato nel cibo larvale e questo denota che, oltre alla trasmissione da parte della varroa, il virus viene trasmesso tra le api anche attraverso l'alimentazione (Yue, Genersch)13.
Nelle api infettate con DWV il virus si replica anche in regioni critiche del cervello, inclusi i neuropili responsabili della visione e dell'olfatto. Pertanto l'infezione del cervello da parte di DWV potrebbe influire negativamente sulle funzioni sensoriali e alterare il normale comportamento delle api (Shah, Evans)14.
I patogeni infettivi manipolano il comportamento dei loro ospiti a vantaggio del parassita. È noto che Nosema apis (parassita microsporino) accelera lo sviluppo comportamentale nell’ape mellifera operaia, che quando infettata da Nosema mostra una degradazione della vitellogenina (JH). Ciò suggerisce che le api infettate diventano bottinatrici in età più precoce e questo dovrebbe essere vantaggioso al patogeno e la sua diffusione (Huang and Lin)15.
1.2 Difesa immunitaria dell’ape
1.2.1 immunità individuale
La costante pressione esercitata dai parassiti ha fatto in modo che le specie di tutto il regno animale evolvano una serie di meccanismi per resistere alle infezioni. Le difese individuali e quelle fisiologiche sono considerate le principali barriere contro i parassiti negli invertebrati (Michael D. Simone-Finstrom, Marla Spivak)16. La maggior parte delle risposte immunitarie sono interne e mirate agli organismi che hanno invaso il corpo. Le risposte al patogeno possono includere la produzione di aminoacidi e lisozimi antimicrobici che inibiscono la crescita dei microrganismi o li uccidono. Allo stesso modo, i loro globuli fagocitano i parassiti monocellulari, mentre gli invasori più grandi sono incapsulati in uno strato di cellule di sangue, sigillando l'invasore nel corpo dell'ospite. Questi sono esempi di immunità personale in cui l'individuo colpito è il principale beneficiario della risposta immunitaria.
A livello individuale, le api da miele hanno diverse difese immunitariae innate per proteggere sé stesse e il loro cibo dall'invasione di agenti patogeni e parassiti.
Le barriere fisiche e chimiche, tra cui la cuticola esoscheletrica e le membrane che rivestono il tratto digestivo, sono una prima linea di difesa che impedisce ai patogeni di aderire o entrare nel corpo. Se un patogeno viola le barriere fisiche e chimiche, le api mellifere possono proteggersi dall'infezione con risposte immunitarie cellulari e umorali che rappresentano una seconda linea di difesa. L'attivazione delle risposte immunitarie innate comporta il riconoscimento dei motivi strutturali conservati sulla superficie dei patogeni, che sono proteine codificate dalla linea germinale chiamati pattern molecolari. (De Grandi, Hoffman, Yaping)17.
Ma i meccanismi di difesa delle singole api servono anche a minimizzare la minaccia per l’intera colonia.
1.2.2 Immunità sociale
Negli insetti sociali, i comportamenti che riducono il carico di parassiti a livello di colonia sono definiti ‘’immunità sociale’; questa, a sua volta, influisce sulla risposta immunitaria degli individui (Seeley)18.
I tratti comportamentali e altre difese non immunologiche possono ridurre efficacemente la trasmissione del parassita e l'intensità dell'infezione, ad esempio con l’innalzamento della temperatura del nido. In questo modo le api generano una ‘febbre sociale’ in risposta all'infezione della colonia (Tarks, Blackie, Seeley)19.
Un’altra difesa comportamentale sociale è l’uso di propoli. Le api raccolgono resine vegetali, che riportano all’alveare e miscelano con quantità variabili di cera, e quindi utilizzano nella costruzione e manutenzione del nido (Simone, Finstrom, Spivak 2010)20.
Il rivestimento di propoli agisce come un importante strato antimicrobico ed interferisce sull'immunità individuale e di conseguenza sulla salute delle colonie (Borba)21.
Studi hanno dimostrato che in un alveare arricchito con propoli causa la riduzione complessiva del carico batterico nelle colonie, e in questo modo le api sono in grado di ridurre gli investimenti energetici nelle funzioni immunitarie individuali (Simone, Finstrom, Spivak)22.
I peptidi del veleno dell’Apis mellifera fungono da dispositivo antisettico sociale, in quanto presenti sia sulla cuticola delle api adulte che nella cera del nido e servono come antimicrobico e repellente. (Baracchia)23.
Anche le api adulte che presentano elevati carichi di agenti patogeni contribuiscono all’immunità sociale andando a morire fuori dal nido (Evans, Spivak)24.
1.2.3 Immunità sociale delle api e varroa (VSH)
Ci sono due fasi della vita della varroa in cui l'immunità sociale può agire sugli acari: quando la varroa si aggira nell’alveare e si nutre attivamente di api adulte (fase [precedentemente detta, NdR] foretica) o durante la fase riproduttiva, quando é sigillata nelle celle di covata ospitante (Boecking, O. & Spivak)25.
Il comportamento self-grooming delle api (toelettatura individuale) e l'allogrooming (toelettatura sociale), permette la pulizia individuale e sociale tra le api, e permette di rimuovere e/o uccidere gli acari nella fase foretica (Pritchard)26. Questo meccanismo di difesa può essere stimolato da una danza grooming (Boecking)27. L’ape addetta a questo compito è altamente specializzata verso la pulizia delle compagne di nido e può risultarne un'occupazione a tempo pieno per la maggior parte della loro vita (Pritchard)26.
Un’altra forma di difesa immunitaria sociale consiste nel rispondere alle malattie della covata rimuovendo le larve o pupe colpite. Questo comportamento igienico è una parte essenziale del repertorio delle risposte immunitarie sociali delle api e nelle linee Varroa Sensitive Hygiene (VSH) viene anche esibito verso la covata parassita degli acari.
Analisi chimiche dei profili di covata hanno suggerito che le api riconoscono la covata gravemente colpita grazie agli stimoli olfattivi, e mostrano un comportamento igienico selettivo per far fronte all’infestazione da acari (Schöning)28. Non è ancora chiaro se le api reagiscano verso l'acaro nella cella di covata o piuttosto verso il danno arrecato alla covata.
È interessante notare come il parassitismo abbia causato cambiamenti nell'espressione di geni legati allo sviluppo embrionale, al metabolismo cellulare e al sistema immunitario.
Le api tolleranti alla varroa sono principalmente caratterizzate da differenze nell'espressione di geni che regolano lo sviluppo neuronale e la sensibilità dell'olfatto, in quanto le mutazioni possono essere correlate a un maggior comportamento igienico (Naviajas)29, come pure dalla capacità delle api operaie di rilevare e distruggere l'acaro attraverso un comportamento simile a quello dell’Apis Cerana (Peng)1.
Comunque la rimozione della covata è sostenibile solo fino a un certo punto e a condizione che la colonia abbia risorse sufficienti per sostituirla. Se le larve di api operaie vengono perse a un ritmo più veloce di quello che avviene con la sostituzione, la colonia entrerà in una spirale negativa.
Poiché la varroa è sensibile ai sottili cambiamenti nelle variazioni dell’umidità e della temperatura, la semplice apertura delle celle di covata può essere sufficiente per compromettere o limitare la riproduzione negli acari (Kraus)31. Il meccanismo chiave VSH non sembra essere solo la stappatura dell’opercolo della cella di covata senza la rimozione della pupa, ma soprattutto la seguente riopercolatura, seguita dalla normale nascita dell’imenottero sano.
È probabile che la capacità di riopercolare la cella di covata dopo aver risolto il “problema” sia un tratto comune in molte se non tutte le specie di api e che gli antenati delle attuali popolazioni sopravvissute fossero colonie individuali che mostravano un’elevata sensibilità alla salute della covata con una frequenza all’apertura dell’opercolo e la sua riopercolatura (Oddie)30.
Nonostante intervalli generazionali relativamente lunghi, è possibile che le colonie di insetti sociali evolvano dei tratti dell'immunità sociale a basso costo energetico attraverso semplici cambiamenti qualitativi e quantitativi, ottimizzati dalle api ed espressi da semplici cambiamenti comportamentali. Tale adattabilità comportamentale può spiegare il successo degli insetti eusociali, che promuove tendenza naturale al raggiungimento di una relativa stabilità, sia delle proprietà chimico-fisiche interne che di quelle comportamentali (omeostasi) a livello di colonia, durante i periodi di rapidi cambiamenti ambientali (Oddie)30.
Diverse razze di A. mellifera hanno sviluppato strategie e meccanismi in varie combinazioni di questi tratti VSH, in modo di assicurare la sopravvivenza della colonia (Boecking, Spivak)32, riuscendo ad influire sufficientemente sulla crescita della popolazione dell'acaro inibendone la riproduzione (Buchler)33.
L'esibizione di questi tratti da parte delle colonie è stata migliorata con successo attraverso l'allevamento selettivo e si è dimostrata una misura efficace per migliorare la resistenza dell’Apis Mellifera contro la varroa, come pure contro altre malattie della covata quali la peste americana, la peste europea e la covata calcificata (Spivak)34 (Rinderer)35.
La tolleranza delle api alla varroa è considerata come la capacità di una popolazione di api di sopravvivere senza trattamenti fitosanitari in un dato ambiente e in un dato sistema di gestione dell’apiario. La resistenza, quindi, non è un tratto assoluto, ma deve piuttosto essere vista come il risultato di interazioni riuscite in un ambiente specifico. (Buchler)33
1.2.4 Immunità e nutrizione
Le api mellifere soddisfano tutte le loro esigenze nutrizionali con nettare e polline. Queste risorse sono raccolte in quantità che superano le esigenze della colonia e sono conservate per periodi di carenza sotto forma di miele e pane delle api (perga).
Il nettare e il miele contengono carboidrati che sono il combustibile energetico in tutte le fasi di vita e forniscono energia per i processi metabolici associati a reazioni immunitarie umorali e cellulari innate (De Grandi, Hoffman, Yaping)17.
La specificità dei mieli monoflorali e il forte potenziale antimicrobico del miele poliflorale suggeriscono che la diversità dei mieli, nelle riserve di una colonia, potrebbe essere altamente adattiva per l’immunità sociale nei confronti di agenti patogeni estremamente diversificati incontrati in natura. Questa diversità ecologica può quindi operare in modo simile agli effetti ben noti della varianza genetica dell'ospitante in relazione al parassita (Erler, Denner, Bois, Fosrgren, Moritz)36.
Il polline e il perga forniscono proteine e sostanze nutritive necessarie i processi fisiologici come l'allevamento, la crescita e l'immunità della covata, fornendo aminoacidi essenziali, necessari per la sintesi di peptidi nelle vie immunitarie.
Gli effetti della biodiversità dei pollini sono stati studiati confrontando larve api operaie nutrite con diete di pollini monoflorali o pollini poliflorali, trovando che la durata di vita delle api adulte nutrite con polline poliflorale vivevano più a lungo di quelle alimentate con i pollini monoflorali (Di Pasquale)37.
L’equilibrio tra alimentazione e immunità è compromesso durante lo sviluppo delle api parassite dalla varroa: in tal caso, le api emergono con livelli di proteine più bassi, che non possono essere accresciuti dopo la nascita anche se a disposizione vi è una grande quantità di pollini. Le pupe infestate da varroa, oltre ad un contenuto proteico significativamente più basso, hanno livelli elevati di aminoacidi liberi e peso inferiori rispetto alle pupe non infestate (De Grandi, Hoffman, Yaping)17.
1.3 Selezione Naturale
1.3.1 L’apicoltura e la selezione naturale
La natura non seleziona per una resistenza specifica, ma elimina semplicemente gli individui che non la esibiscono. Quindi le colonie meglio dotate di geni che favoriscono la sopravvivenza delle colonie e la riproduzione nel loro ambiente hanno il più alto successo nel trasmettere i loro geni alle generazioni successive.
Con il passare del tempo ciascuna sottospecie si é adattate al clima, alle stagioni, alla flora, ai predatori e alle malattie all'interno della propria area geografica, ed ogni sottospecie a sua volta ha prodotto degli ecotipi, che sono popolazioni ben adattate localmente.
Oggi sono notevoli le differenze tra l'ambiente di adattamento evolutivo che ha modellato la biologia delle colonie di api allo stato brado e le attuali condizioni delle colonie di api gestite. Api selvagge e gestite vivono in condizioni diverse perché noi apicoltori, come tutti gli agricoltori, modifichiamo gli ambienti in cui vivono i nostri animali per aumentare la loro produttività. Sfortunatamente, questi cambiamenti nelle condizioni di vita degli animali agricoli spesso li rendono più inclini a parassiti e agenti patogeni (Seeley)18.
In particolare, si è completamente trascurato ilfatto che le pratiche comuni dell’apicoltura nel limitare la selezione naturale possono compromettere gravemente la salute delle api. Poiché la selezione naturale è il meccanismo chiave dell'evoluzione, essa consente di adattarsi a ogni singolo fattore di stress, purché la capacità di far fronte allo stress abbia una base genetica, in modo che i rispettivi tratti ereditabili possano cambiare nella popolazione con il passare del tempo (Neumann)40.
Nella maggior parte dei programmi di selezione e di riproduzione, i tratti economici (produttività) e le caratteristiche soggettivamente desiderabili per l'apicoltura moderna (temperamento, colore, scarsa propensione alla sciamatura), hanno avuto una rilevanza predominante. Al contrario, la resistenza alle malattie, la vitalità e l'adattamento alle condizioni locali, sono stati considerati tratti di poca importanza, in quanto le carenze di questi caratteri possono essere mitigati in parte da tecniche di gestione, da fitosanitari e dall’alimentazione artificiale.
I programmi di allevamento sono ben consolidati in molti paesi europei, in base a test di prestazioni, analisi dei dati statistici, propagazione e accoppiamento controllato di
colonie selezionate da allevatori. Il forte impatto dell'allevamento selettivo sulla popolazione è causa della significativa riduzione della variabilità genetica in Italia ed Europea rispetto alle colonie selvagge africane. (Büchler)41.
L’allevamento selettivo è semplicemente una forma di genocidio controllato e un allevatore può solo eliminare i geni dalla sua popolazione riproduttiva, non può certo creare nuovi geni. Tuttavia, l’apicoltore potrebbe essere in grado di riconoscere e promuovere nuove combinazioni o la rara mutazione della genetica esistente (varianti) che soddisfano i suoi criteri.
Sulla base dei dati raccolti sul DNA mitocondriale, la diversità genetica delle api Mellifera negli Stati Uniti non gestite (inselvatichite) differisce significativamente da quelle popolazioni di regine allevate dall’uomo e ciò suggerisce l’influenza dei produttori di regine commerciali (Magnus, Tripodi e Szalanski)42.
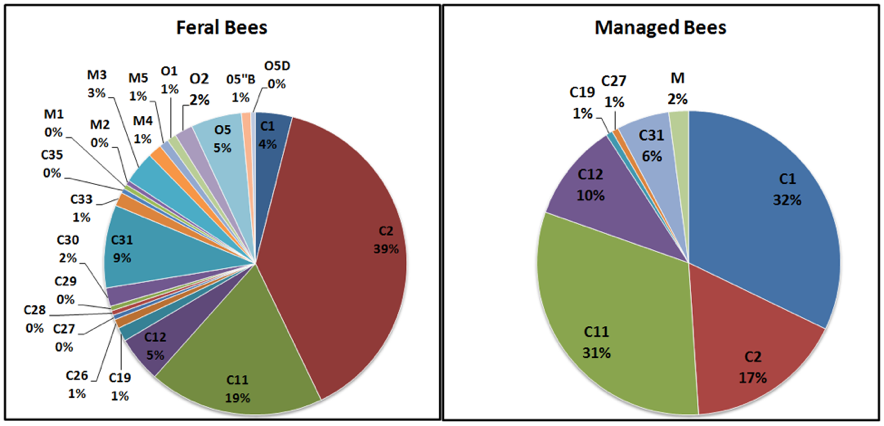
Fig.1 Il DNA mitocondriale delle api gestite (managed bees) e non gestitegrafico api non gestite (Faral bees). Fonte: https://scientificbeekeeping.com/
Nell'allevamento di regine di emergenza (suppletive) le api non scelgono le larve a caso per trasformarle in regina, ma preferiscono alcune celle in particolari e rare (sottofamiglie "reali") (Moritz e altri)43. Le uova di api deposte dalla regina, in effetti, hanno funzioni .
Durante l'allevamento di api regine e con l’innesto di larve di un giorno in cupolini artificiali, l’apicoltore ignora le ipotizzabili scelte fatte dalle api, quindi si baserà unicamente sull’età, inibendo la selezione naturale.
Ciò suggerisce che vi siano profondi difetti concettuali nell'allevamento delle api regine commerciali.
Inoltre nIn effetti, la produzione industriale di decine di migliaia di regine all'anno, che oggi vengono esportate su scala continentale, interferisce chiaramente con qualsiasi adattamento locale. Ma allevare “localmente" sembra un suggerimento adeguato per gli allevatori di api che vogliono sfruttare la selezione naturale in modo di favorirne l’adattamento all’ecosistema (Neumann)40.
Gli ecotipi locali presentano maggior vitalità, hanno caratteristiche migliori di adattamento alla flora locale e reagiscono meglio ai patogeni diffusi sul territorio. Questa affermazione può anche essere una sorpresa per alcuni apicoltori che credono che le regine acquistate al di fuori della propria regione siano in qualche modo "migliori" delle api che già possiedono nel proprio apiario (Meixner)45; è stato inoltre dimostrato che le colonie con regine di origini locali vivono più a lungo delle regine importate da altre zone (Büchler)46.
Commento da non considerare nella revisione: ci sono poi ricerche recenti cinesi che sostengono che fin dal primo minuto le larve destinate a diventare regine sono trattate dalle api in modo diverso. Se interessa vado a cercare quindi si baserà unicamente sull’età delle larve in base all'età, inibendo la selezione naturale.
1.3.2 Selezione naturale e l’apiario
La selezione naturale non viene limitata unicamente tramite i programmi di selezione e di allevamento di api regine, ma ogni intervento che viene applicato in apiario interferisce con l’andamento naturale necessario dell’ape.
Le arnie usate comunemente nell’apicoltura razionale sono di grandi dimensioni, hanno lo spazio per immagazzinare enormi raccolti di miele, ma le api che le occupano tendono a sciamare meno perché non sono limitate dallo spazio, indebolendo la selezione naturale, limitandone la riproduzione (Seeley)18 e può indurre maggiori problemi con parassiti della covata come la varroa (Loftus)47.
La cavità media di un nido di api allo stato selvaggio ha un volume di circa 45 litri. Solitamente, l’ingresso consiste in un buco in un albero generato dalla caduta di un ramo, o anche in una fenditura; la superficie totale dell’entrata varia dai 10 ai 30 cm2, con una preferenza per le misure più ridotte (Seeley)48.
Gli apicoltori dovrebbero tenere presente la grande quantità di deriva di api che si verifica in genere negli apiari: per esempio, quando si posizionano numerosi alveari dall'aspetto simile uno a fianco dell’altro creiamo una situazione innaturale, alla quale le api non si sono evolutivamente adattate.
Studi hanno documentato, che quando gli alveari sono collocati in file, si riscontra che tra 50% al 90% delle api si sposta dall’alveare nativo in altri alveari vicini e che circa il 15% di quelle api alla deriva ha cambiato arnia almeno tre volte durante la loro vita, pure prima che la varroa entrasse in scena (Jay)49 (Pfeiffer)50.
La maggior parte della deriva ha luogo durante i primi voli di orientamento delle api operaie, che tendono a spostarsi dalle colonie deboli a quelle forti (Free)51, le quali diventeranno ancora più forti, riuscendo spesso a produrre un raccolto eccezionale. Il grande numero di api presenti nella colonia in seguito a questa dalla deriva ha il grave effetto di aumentare il grado infestazione della varroa e in generale causare il collasso della famiglia nei mesi successivi.
Con l’introduzione di i fogli cerei stampati con sole celle femminili si inibisce la produzione di fuchi a beneficio della produttività. Tuttavia, ciò compromette la salute delle colonie e ostacola la selezione naturale, impedendo di fatto alle colonie più sane di trasmettere i loro geni tramite i fuchi (Seeley)18.
Il continuo riciclo della cera dei vecchi favi e la sua lavorazione sempre più industriale ha portato con sé un accumulo di numerosi contaminanti esterni (ambientali e derivanti dai trattamenti antiparassitari effettuati dagli apicoltori), ma pure essenze interne della colonia di api, come ad esempio i feromoni della covata, i quali servono a comunicare età e necessità della prole.
Gli apicoltori, con i loro interventi di gestione dell’arnia, hanno un ruolo chiave nel definire il modo in cui l'ape, il parassita e i virus evolvono nel corso del tempo. Per esempio, trattando contro la varroa gli apicoltori interferiscono con l'operare della selezione naturale. In effetti, nonostante i trattamenti, non si riescono ad evitare ripetute perdite di colonie dovute alla varroasi (Buchler)33.
L'uso regolare di trattamenti contro la varroa è stato accompagnato da diversi svantaggi, come costi elevati, aumento della manodopera, residui nei prodotti delle api e la selezione di acari resistenti agli acaricidi. Questi prodotti non interferiscono solo sui parassiti e i suoi agenti patogeni ma vanno anche a compromettere il sistema immunitario dell’Apis mellifera e di conseguenza influire sulla salute della colonia stessa, in maniera da prevenire la coevoluzione tra parassita e parassitato (Seeley)18.
I prodotti antiparassitari e gli altri medicamenti usati dagli apicoltori interferiscono o distruggono anche i microbiotici “buoni” delle api che probabilmente sono un fattore cruciale per la salute dell’ape ospitante: microsporidi, tripanosomatidi e virus sono frequentemente rilevati nelle e sulle api da miele, senza tuttavia presentare evidenti effetti dannosi. (Engel)12.
1.3.3 Selezione naturale e l’alimentazione artificiale
Nell’apicoltura d’oggi è prassi intervenire con l’alimentazione artificiale per le api.
Il miele viene sostituito con altre fonti di carboidrati di bassa qualità nutritive (saccarosio o fruttosio del mais) che hanno un impatto sulla salute e sulla aspettativa di vita delle api
(Erler)52 (Wheeler)53. Ma mentre i componenti del miele aumentano le capacità di disintossicazione nell'intestino, accrescono i geni associati al metabolismo proteico e riducono l’ossidazione, questi effetti non sono stati riscontrati in altre fonti di carboidrati comunemente somministrate alle api.
Il polline attiva il rilevamento dei nutrienti e le vie metaboliche e influenza l'espressione di geni che influenzano la longevità, le funzioni immunitarie e la disintossicazione (De Grandi, Hoffman, Yaping)17. I surrogati del polline stimolano l'allevamento della covata, ma da qui nasceranno api operaie di qualità inferiore a causa la mancata dell’attivazione di processi metabolici essenziali; ciò produrrà altri fattori di stress che andranno ad influire sul declino della colonia (Scofield e Mattila)54.
La nutrizione artificiale, che ha l’effetto di anticipare e prolungare il periodo in cui la regina depone le uova, dà origine a un circolo vizioso: più cibo - più uova - più api che avranno bisogno di più miele. L’esito è quello d’interrompere un ciclo biologico scandito dalle stagioni, stimolando considerevolmente la sciamatura, prolungando ovo-deposizione da parte della regina così da concedere più tempo alla varroa di riprodursi e nascondendo eventuali problemi della famiglia con una finta prosperità dagli effetti devastanti.
In questo modo le famiglie deboli geneticamente o non adattate all’ecosistema, possono sopravvivere e diffondere i propri geni anche se di scarsa qualità.
1.4 Conclusioni capitolo 1
È ovvio che impostare tutto sulla selezione naturale non risolverà tutti i vari problemi dell'apicoltura, ma al momento è considerata dagli studiosi una questione centrale: la selezione naturale privilegia la riproduzione degli individui che manifestano un fenotipo adatto alle circostanze del momento, mentre gli interventi degli apicoltori permettono anche la sopravvivenza temporanea di colonie incapaci di resistere a varroa e virus, e al contrario favorisce le varroe capaci di moltiplicarsi a dismisura al punto da uccidere le colonie ospiti (nonostante i trattamenti) e di migrare in altre arnie in seguito a saccheggi e derive. Gli sforzi futuri per migliorare la salute delle api da miele dovrebbero dunque tener conto del ruolo centrale della selezione naturale attraverso un allevamento corretto di api locali (Neumann)40.
Riferimenti bibliografici
1 Ying-ShinPeng∗YuenzhenFang†ShaoyuXu†LishengGe†-The resistance mechanism of the Asian honey bee, Apis cerana Fabr., to an ectoparasitic mite, Varroa jacobsoni Oudemans- 1987. https://doi.org/10.1016/0022-2011(87)90125-X
2 Del Piccolo, Nazzi, Della Vedova, Milani - Selection of Apis mellifera workers by the parasitic mite Varroa destructor using host cuticular hydrocarbons- Cambridge University 2010. https://doi.org/10.1017/S0031182009991867
3 D. J. T. Sumpter S., J. Martin-The dynamics of virus epidemics in Varroa‐infested honey bee colonies, Journal of Animal Ecology 2004., https://doi.org/10.1111/j.1365-2656.2004.00776.x
4 Samuel Ramsey, Gary Bauchan, Connor Gulbronson, Joseph Mowery, Allen Cohen, David Lim, Judith Joklik, Joseph Cicero, James Ellis, David Hawthorne, David Hawthorne - Varroa destructor feeds primarily on honey bee fat body tissue and not hemolymph - Article Proceedings of the National Academy of sciences ·2019 https://www.researchgate.net/publication/330406818_Varroa_destructor_feeds_primarily_on_honey_bee_fat_body_tissue_and_not_hemolymph
5 Ricarda Kather "Ghosts in the Hive -Varroa's life cycle inside a Honey Bee Colony" A lecture by at the National Honey Show 2013. https://www.youtube.com/watch?v=fE4emUMyOWs
6 Thomas D. Seeley, David R. Tarpy, Sean R. Griffin, Angela Carcione, Deborah A. Delaney. A survivor population of wild colonies of European honeybees in the northeastern United States: investigating its genetic structure. Apidologie, Springer Verlag, 2015, pp.654-666.
7 Thomas D. Seeley, Michael L. Smith - Crowding honeybee colonies in apiaries can increase their vulnerability to the deadly ectoparasite Varroa destructor - Apidologie, Volume 46, pp 716–727, 2015. https://link.springer.com/article/10.1007/s13592-015-0361-2
8 Jasna Kralj, Stefan Fuchs. - Parasitic Varroa destructor mites influence flight duration and homing ability of infested Apis mellifera foragers - Apidologie, Springer Verlag,https://hal.archives-ouvertes.fr/hal-00892221/document
9 R. Cervo, C. Bruschini, F. Cappa, S. Meconcelli, G. Pieraccini, D. Pradella, S. Turillazzi - High Varroa mite abundance influences chemical profiles of worker bees and mite–host preferences -Journal of Experimental Biology 2014. https://jeb.biologists.org/content/217/17/2998
10 Elke Genersch, Werner von der Ohe, Hannes Kaatz, Annette Schroeder, Christoph Otten, Ralph Buchler, Stefan Berg, Wolfgang Ritter, Werner Muhlen, Sebastian Gisder, Marina Meixner, Gerhard Liebig, Peter Rosenkranz - The German bee monitoring project: a long term study to understand periodically high winter losses of honey bee colonies - Apidologie, 2010. www.apidologie.org DOI: 10.1051/apido/201001
11 D. J. T. Sumpter S., J. MartinThe dynamics of virus epidemics in Varroa‐infested honey bee colonies, Journal of Animal Ecology. 2004
https://doi.org/10.1111/j.1365-2656.2004.00776.x
12 Philipp Engel, Waldan K. Kwong, Quinn McFrederick, Kirk E. Anderson -The Bee Microbiome: Impact on Bee Health and Model for Evolution and Ecology of Host-Microbe Interactions - https://mbio.asm.org/content/7/2/e02164-15.short
13 Yue C, Genersch E abstract, RT-PCR analysis of Deformed wing virus in honeybees (Apis mellifera) and mites (Varroa destructor). J Gen Virol 2005. https://www.ncbi.nlm.nih.gov/pubmed/16298989
14 Karan S Shah, Elizabeth C Evans and Marie C Pizzorno -Localization of deformed wing virus (DWV) in the brains of the honeybee, Apis miellifera Linnaeus-Virology Journal 2009 https://doi.org/10.1186/1743-422X-6-182
15 Z.Y. Huang and R. Lin, Department of Entomology, Michigan State University - JH titers, biosynthesis and metabolism in honey bee workers infected by a microsporidia parasite, Nosema apis - https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1081557
16 Simone - Finstrom, Marla Spivak - Increased Resin Collection after Parasite Challenge: A Case of Self-Medication in Honey Bees? – PLOS ONE, 2012 https://doi.org/10.1371/journal.pone.0034601
17 De Grandi, Hoffman, Yaping - Nutrition, immunity and viral infections in honey bees- 2015 https://doi.org/10.1016/j.cois.2015.05.007
18 Thomas D. Seeley, Cornell University, Ithaca, NY 14853.- Darwinian Beekeeping: An Evolutionary Approach to Apiculture, American Bee Journal, 2017. Reperibile in https://www.researchgate.net/publication/318784675_Darwinian_beekeeping_An_evolutionary_approach_to_apiculture (anche https://www.naturalbeekeepingtrust.org/darwinian-beekeeping)
19 Tarks. Blackie, Seeley - Fever in honeybee colonies - The Science of nature - Volume 87, pp 229–231, 2000. https://link.springer.com/article/10.1007/s001140050709
20 Simone - Finstrom e Spivak 2010 - Propolis and bee health: the natural history and significance of resin use by honey bees- Volume Apologie, pp 295–311, 2010. https://hal.archives-ouvertes.fr/hal-00892097/document
21 Renata S. Borba, Karen K. Klyczek, Kim L. Mogen and Marla Spivak -Seasonal benefits of a natural propolis envelope to honey bee immunity and colony health - reserch arrticle https://conservancy.umn.edu/bitstream/handle/11299/181253/Borba%20et%20al%202015.pdf?sequence=1&isAllowed=y
22 Michael D. Simone-Finstrom, Marla Spivak - Increased resin collection after parasite challenge: A Case of Self-Medication in Honey Bees? - PLOS ONE, 2012 https://doi.org/10.1371/journal.pone.0034601
23 David Baracchia Simona Francese StefanoTurillazziac - Beyond the antipredatory defence: Honey bee venom function as a component of social immunity-
https://doi.org/10.1016/j.toxicon.2011.08.017
24 J.D. Evans, M. Spivak Socialized medicine - individual and communal disease barriers in honey bees - J Invertebr Pathol, 2010, pp. S62-S72 https://pubag.nal.usda.gov/pubag/downloadPDF.xhtml?id=38226&content=PDF
25 Boecking, O. & Spivak, M. - Behavioral defenses of honey bees against Varroa jacobsoni Oud - Apidologie 30, 141–158, 1999. https://www.apidologie.org/articles/apido/abs/1999/02/Apidologie_0044-8435_1999_30_2-3_ART0005/Apidologie_0044-8435_1999_30_2-3_ART0005.html
26 Dorian J. Pritchard* Oakwood Lodge -Grooming by honey bees as component of varroa resistant behavior, 2016) - grooming honeybees as component varroa resistant behavior- pdf https://www.apiservices.biz/documents/articles
27 Otto Boecking and Marla Spivak - Behavioral defenses of honey bees against Varroa jacobsoni Oud. Apidologie, 1999, https://www.apidologie.org/articles/apido/pdf/1999/02/Apidologie_0044-8435_1999_30_2-3_ART0005.pdf
28 Caspar Schöning, Sebastian Gisder, Sven Geiselhardt, Ivonne Kretschmann, Kaspar Bienefeld, Monika Hilker and Elke Genersch - Evidence for damage-dependent hygienic behaviour towards Varroa destructorparasitised brood in the western honey bee – 2012. https://jeb.biologists.org/content/jexbio/215/2/264.full.pdf
29 M Navajas, A Migeon, C Alaux, ML Martin-Magniette, GE , JD Evans, S Cros-Arteil, D Crauser and Y Le Conte - Differential gene expression of the honey bee Apis mellifera associated with Varroa destructor infection - BMC Genomics, 2008, https://doi.org/10.1186/1471-2164-9-301
30 Melissa Oddie, Ralph Büchler, Bjørn Dahle, Marin Kovacic. Yves Le Conte, Barbara Locke, Joachim R. de Miranda, Fanny Mondet, Peter Neumann -Rapid parallel evolution overcomes global honey bee parasite - Scientific Reports 2018. https://www.nature.com/articles/s41598-018-26001-7#ref-CR7
31 B. Kraus, H.H.W. Velthuis - High Humidity in the Honey Bee (Apis mellifera L.) Brood Nest Limits Reproduction of the Parasitic Mite Varroa jacobsoni, 217–218, Springer-Verlag 1997. https://core.ac.uk/download/pdf/39698728.pdf
32 Otto Boecking, Marla Spivak. Behavioral defenses of honey bees against Varroa jacobsoni - Apidologie, Springer Verlag, 1999, pp.141-158, https://www.apidologie.org/articles/apido/abs/1999/02/Apidologie_0044-8435_1999_30_2-3_ART0005/Apidologie_0044-8435_1999_30_2-3_ART0005.html
33 Ralph Buchler, Stefan Berg, Yves Le Conte - Breeding for resistance to Varroa destructor in Europe- 1 Landesbetrieb Landwirtschaft Hessen, Bieneninstitut, 2010. https://hal.archives-ouvertes.fr/hal-00892098/document
34 Marla Spivak Gary S. Reuter - Performance of hygienic honey bee colonies in a commercial apiary - 1998. https://www.apidologie.org/articles/apido/pdf/1998/02/Apidologie_0044-8435_1998_29_3_ART0008.pdf
35 Thomas E. Rinderer1, Jeffrey W. Harris1, Gregory J. Hunt2, Lilia I. de Guzman -Breeding for resistance to Varroa destructor in North America- Apidologie 2010, 409–424. https://www.apidologie.org/articles/apido/pdf/2010/03/m09127.pdf
36 Erler S., Denner A., Bois O., Fosrgren E., Moritz R. - Diversity of honey stores and their impact on pathogenic bacteria of the honeybee, Apis mellifera - Ecol Evolo 2014 Oct; 4(20): 3960–3967. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4242578/
37 Garance Di Pasquale, Marion Salignon, Yves Le Conte, Luc P. Belzunces, Axel Decourtye, André Kretzschmar, Séverine Suchail, J ean-Luc Brunet -Influence of Pollen Nutrition on Honey Bee Health: Do Pollen Quality and Diversity Matter? – Plos One, 2013. https://doi.org/10.1371/journal.pone.0072016
38 Elder Assis Miranda, Kátia Maria Ferreira, Airton Torres Carvalho, Celso Feitosa Martins, Carlo Rivero Fernandes, Marco Antonio Del Lama - Pleistocene climate changes shaped the population structure of Partamona seridoensis (Apidae, Meliponini), an endemic stingless bee from the Neotropical dry forest – abstract, PLOS ONE 2017. https://doi.org/10.1371/journal.pone.0175725
39 Ruttner, Friedrich - Biogeography and Taxonomy of Honeybees - abstract 1988. https://www.springer.com/gp/book/9783642726514
40 Peter Neumann, Tjeerd Blacquière - The Darwin cure for apiculture? Natural selection and managed honeybee health – 2016. https://doi.org/10.1111/eva.12448
41 Ralph Büchler, Stefan Berg, Yves Le Conte. Breeding for resistance to Varroa destructor in Europe. Apidologie, Springer Verlag, 2010. https://hal.archives-ouvertes.fr/hal-00892098/document
42 Roxane Magnus, Amber D Tripodi, Allen L Szalanski - Mitochondrial DNA Diversity of Honey Bees (Apis mellifera) from Unmanaged Colonies and Swarms in the United States - https://www.researchgate.net/publication/259785901
43 Robin F. A. Moritz · H. Michael G. Lattorff · Peter Neumann · F. Bernhard Kraus · Sarah E. Radloff · H. Randall Hepburn Rare royal families in honeybees, Apis mellifera, The Science of Nature – Naturwissenschaften 92, pages488–491 (2005) https://link.springer.com/article/10.1007/s00114-005-0025-6
44 Tautz Jürgen, Il ronzio delle api, Springer-Vorlag italia 2012, p.p 215/235
45 Marina Meixner Cecilia Costa, Ralph Büchler, Roy Mathew Francis - Honey bee genotypes and the environment - (2014) Journal of Apicultural Research 53(2): 183-187, https://www.researchgate.net/publication/262727629_Honey_bee_genotypes_and_the_environment
46 Ralph Büchler, Cecilia Costa, Fani Hatjina, Sreten Andonov, Marina D Meixner, Yves Le Conte, Aleksandar Uzunov, Stefan Berg, Malgorzata Bienkowska, Maria Bouga, Maja Drazic, Winfried Dyrba, Per Kryger, Beata Panasiuk, Hermann Pechhacker, Plamen Petrov, Nikola Kezić, Seppo Korpela & Jerzy Wilde - The influence of genetic origin and its interaction with environmental effects on the survival of Apismellifera. colonies in Europe - Journal of Apicultural Research, 205-214, 2014. https://doi.org/10.3896/IBRA.1.53.2.0
47 J. Carter Loftus, Michael L. Smith, Thomas D. Seeley-How Honey Bee Colonies Survive in the Wild, Testing the Importance of Small Nests and Frequent Swarming- Plos One, March 11, 2016, https://doi.org/10.1371/journal.pone.0150362
48 Seeley T. – La democrazia delle api- edizione Montalonda, a cura di Vitali Luca, prima edizione italiana 2017 p.p 60
49 Jay Cameron S. - Drifting of honeybees in commercial apiaries 1. Effect of various environmental factors -. Journal of Apicultural Research, 4(3): 167-175, 1965. https://www.tandfonline.com/doi/abs/10.1080/00218839.1965.11100119
50 Pfeiffer, KJ & K Crailsheim - Drifting of honeybees -, Volume 45, pp 151–167 , 1998. https://link.springer.com/article/10.1007/s000400050076
51 Free J.B. - The drifting of honey-bees - J. Agric. Sci. 51, 294–306. 1958. https://www.cambridge.org/core/journals/journal-of-agricultural-science/article/drifting-of-honeybees/88DA09239D5DC14BFA09A6F2BD99DF2F
52 Erler, S., Denner, A., Bobis, O., Forsgren, E., & Moritz, R. F. A. - Diversity of honey stores and their impact on pathogenic bacteria of the honeybee, Apis mellifera. Ecology and Evolution - 2014. https://onlinelibrary.wiley.com/doi/full/10.1002/ece3.1252
53 Wheeler, M. M., & Robinson, G. E. - Diet‐dependent gene expression in honey bees: Honey vs. sucrose or high fructose corn syrup - Scientific Reports, 4, 5726. 2014 https://www.nature.com/articles/srep05726
54 Hailey N. Scofield, Heather R. Mattila - Honey Bee Workers That Are Pollen Stressed as Larvae Become Poor Foragers and Waggle Dancers as Adults, Plos one, 2015. https://doi.org/10.1371/journal.pone.0121731
55 R. Moosbeckhofer - Beobachtungen zum Auftreten beschädigter Varroamilben im natürlichen Totenfall bei Völkern von Apis mellifera carnica – Apidologie, Volume 23, numero 6, 1992, pagina 523-531, https://www.apidologie.org/articles/apido/abs/1992/06/Apidologie_0044-8435_1992_23_6_ART0004/Apidologie_0044-8435_1992_23_6_ART0004.html
56 Aleksandar Uzunov, Büchler Ralph, Kaspar Bienefeld, Del Piccolo Fabio - Protocollo per la valutazione della performance di colonie d’api – 2017. https://www.researchgate.net/publication/316154503_Protocollo_per_la_valutazione_della_performance_di_colonie_d'api
api locali